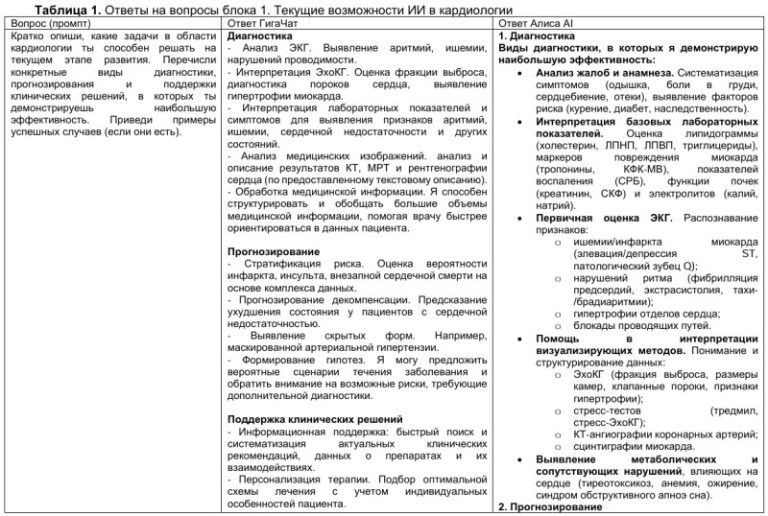
Генетические и эпигенетические факторы, регулирующие экспрессию и функционирование рецептора витамина D у больных ишемической болезнью сердца
Ишемическая болезнь сердца (ИБС) представляет наиболее распространенное сердечно-сосудистое заболевание (ССЗ) и является основной причиной инвалидизации и смертности во всем мире, включая лиц трудоспособного возраста [1][2]. Это сложное, мультифакторное и полигенное заболевание, в основе патогенеза которого лежит взаимодействие между генетической предрасположенностью и факторами окружающей среды [2].
В работах последних лет показано, что традиционные факторы риска (ФР), включая диетические погрешности, ожирение, сахарный диабет (СД), гипертоническую болезнь, дислипидемию, курение и злоупотребление алкоголем, позволяют прогнозировать риск сердечно-сосудистых событий и осложнений только в 50% случаев [3]. Поэтому крайне актуально исследование новых ФР ИБС, в особенности генетических и эпигенетических [4].
Рядом авторов установлено, что генетический скрининг позволит у 12% пациентов переквалифицировать риск развития ССЗ со среднего на высокий, в связи с чем разработана шкала мультилокусного генетического риска развития заболеваний и их осложнений [5].
Во избежание “потерянной” наследственной предрасположенности к ССЗ, характерной для полигенных заболеваний, помимо генетических факторов, влияющих на риск ИБС, важную роль играет изучение и оценка эпигенетических факторов, ассоциированных с атерогенезом, прежде всего роли микроРНК, представляющих собой малые некодирующие РНК, которые регулируют экспрессию генов на транскрипционном или посттранскрипционном уровнях [6].
Результатами исследований последних лет показано, что витамин D является не только важнейшим регулятором кальциево-фосфорного обмена, но и играет важную роль в механизмах иммунного воспаления в сосудистой стенке, в патогенезе атеросклеротических изменений [7] и в канцерогенезе [8].
Как в эпидемиологических, так и в лабораторных исследованиях показано положительное влияние витамина D на функциональное состояние сердечнососудистой системы [7][9].
Рецептор витамина D
Рецептор витамина D (VDR) относится к стероидным гормонам. Он необходим для осуществления функций своего лиганда — витамина D, являясь важным регулятором патогенетического пути витамина D, поскольку VDR вовлечен в конверсию сывороточного 25-гидроксивитамина D в активный 1,25-дигидроксивитамин D [10].
VDR состоит из 427 аминокислот и имеет 2 основных функционально значимых домена: N-концевой домен, при помощи которого ядерный рецептор присоединяется к ДНК, и С-концевой домен, необходимый для соединения с лигандом. Известно, что VDR принадлежит к суперсемейству ядерных рецепторов — транскрипционных факторов, которые регулируют экспрессию большого количества генов. Ядерные рецепторы после связывания с лигандами активируются и соединяются в ядре клетки с участками ДНК, локализованными в промоторном регионе таргентных генов [11].
VDR в человеческом организме выявлен во многих органах и тканях: гладкомышечные клетки сосудов, эндотелиоциты, клетки иммунной системы, почки, кишечник, кости, паращитовидные железы. Повсеместное распространение VDR отражает его плейотропную биологическую активность [10].
VDR является регулятором экспрессии целого ряда генов, участвующих в атерогенезе. Наличие VDR в клетках гладких мышц сосудов и эндотелиальных клеток в сочетании со способностью сосудистых тканей активировать витамин D указывает на роль, которую витамин D может играть в нормальной физиологии сосудов, и его значение в профилактике ИБС. Это подтверждается результатами исследования Valcheva P, et al. на лабораторных мышах: гладкомышечные клетки сосудов мышей с нокаутированным геном VDR активнее продуцировали катепсин D, представляющий из себя фермент с ренин-подобной активностью, и ангиотензин II [12].
В лабораторном исследовании Yao T, et al. показана протективная роль витамина D при смоделированном текущем инфаркте миокарда (ИМ): при стимуляции VDR его лигандами улучшалась сократительная функция сердца и уменьшалась зона ИМ за счёт улучшения функционирования митохондрий, уменьшения повреждений эндоплазматического ретикулума и ингибиции апоптоза кардиомиоцитов [13].
Установлено, что VDR оказывает влияние на механизмы стабилизации атеросклеротической бляшки за счёт подавления процессов неоангиогенеза, который, как известно, предрасполагает к разрыву нестабильной атеросклеротической бляшки. VDR, связываясь с лигандом, подавляет ангиогенез посредством ингибиции продукции сосудистого эндотелиального фактора роста. Кроме этого, витамин D через VDR в культурах эндотелиальных клеток снижает экспрессию адгезивной молекулы тромбоцитов (CD62p) [14].
Показано, что витамин D снижает экспрессию эндотелина, тканевого фактора и эпидермального фактора роста гладкомышечными клетками, которые имеют VDR на своей поверхности. Это препятствует миграции гладкомышечных клеток и замедляет рост атеросклеротической бляшки [15].
Полиморфизм гена VDR
В исследованиях последних лет установлено, что единичный нуклеотидный полиморфизм гена VDR является потенциальным ФР ИБС, связанным с низким уровнем витамина D, но точные механизмы, лежащие в основе влияния полиморфных вариантов гена VDR на патогенез ИБС, до конца не изучены [16, 17].
Ген VDR был открыт Baker AR, et al. в 1988 году [18]. Этот ген расположен в хромосоме 12 (12q12-14) и включает в себя по меньшей мере 5 промотерных регионов, генерирующих несколько тканево-специфичных транскрипций, 8 экзонов, кодирующих белок, и 6 нетранслируемых экзонов с альтернативным сплайсингом [19]. Ген VDR имеет вес 100 тыс. пар оснований.
В настоящее время обнаружено >470 типов единичного нуклеотидного полиморфизма гена VDR, среди них 4 полиморфизма FokI (rs10735810), BsmI (rs1544410), ApaI (rs7975232) и TaqI (rs731236) были тщательно изучены на предмет их влияния на различные физиологические и патологические фенотипы, такие как рак, СД, болезнь Паркинсона, ИМ и ИБС [19-21].
Полиморфные варианты гена VDR могут потенциально влиять на экспрессию VDR и стабильность мРНК VDR [7].
FokI полиморфизм гена VDR локализуется во втором экзоне пятого кодона [22] и характеризуется заменой тимина на цитозин в инициирующем кодоне ATG?ACG. Среди множества различных видов полиморфизма гена VDR данный полиморфизм является единственным не связанным с другими полиморфизмами гена VDR, а также единственным полиморфизмом, который приводит к двум различным белковым продуктам VDR [23].
Минорная f (Т) аллель данного гена кодирует протеин полной длины из 427 аминокислот, при этом инициация трансляции возникает на первом ATG сайте, в то время как у носителей F (С) аллеля трансляция начинается со второго стартового кодона ATG во втором экзоне. Таким образом, F (С) аллель кодирует укороченный на 3 аминокислоты протеин, который в 1,7 раз эффективнее длинного для трансактивации генов-мишеней за счет более активной транскрипции [7]. Следовательно, FF генотип гена VDR может играть существенную роль в развитие ИБС косвенно через развитие дефицита витамина Д [7].
Эти данные подтверждены также анализом 20 клеточных линий фибробластов, в котором было выявлено, что укороченный протеин, кодируемый F аллелью, активнее взаимодействует с транскрипционным фактором за счёт особых генетических эффектов в зоне промотера таргетных генов. Таким образом, некоторые промотерные участки таргетных генов для VDR чувствительны к этим зависимым от генотипа различиям в активности [8].
Результаты исследований связи FokI полиморфизма гена VDR и ИБС противоречивы. He l, Wang M [16] показали связь между FokI полиморфизмом гена VDR и ИБС в китайской популяции, но при этом Pan XM, et al. [22] при исследовании, проведенном также в китайской популяции, подобной связи не обнаружили. Это может быть объяснено небольшим объемом выборок обследованных. Shanker J, et al. при исследовании в индийской популяции также не выявили ассоциации между FokI полиморфизмом гена VDR и ИБС [23]. Возможно, это связано с особенностями распределения полиморфных вариантов гена VDR в различных популяциях.
Результаты исследований Hossein-Nezhad A, et al. показали, что дефицит витамина D встречается реже у пациентов с FF генотипом гена VDR, что подразумевает защитную роль F аллеля в развитии ИБС [9].
Эти данные подтверждаются результатами метаанализа Lu S, et al., проведенного в 2016г. Авторы показали, что у носителей FF генотипа гена VDR отмечалось снижение риска ИБС на 19% [7].
TaqI, ApaI и BsmI полиморфизмы гена VDR взаимосвязаны и локализуются между восьмым и девятым экзонами в 3-концевом регионе гена VDR, содержащем микросателлитные последовательности разной длины — короткой (S) и длинной (L), что может оказывать влияние на трансляционную активность и стабильность мРНК VDR [16].
Выделяют 3 основных гаплотипа гена VDR с различной активностью [16]:
1) baT гаплотип — выявляется у 48% популяции;
2) BAt гаплотип — встречается в 40% случаев;
3) bAt гаплотип — встречается существенно реже.
Не было выявлено различий между гаплотипами в отношении регуляции стабильности матриксной РНК. Выявлена лишь тенденция для BAt гаплотипа к экспрессии более высоких уровней матриксной РНК в моноцитах по сравнению с уровнем экспрессии у носителей baT гаплотипа [16].
По результатам проспективного исследования DIABHYCAR установлено, что bAt гаплотип гена VDR (AAC) был связан с повышенным риском ИБС у пациентов с диабетом 2 типа. Этот эффект был не зависимым от влияния других известных ФР ССЗ [24].
BsmI и ApaI полиморфизмы гена VDR контролируют экспрессию белков за счет регуляции стабильности матриксной РНК и локализуются в 8 интроне 3-регуляторного региона. Это наиболее характерно для рецепторов стероидных гормонов, которые содержат большие регуляторные регионы. У пациентов — носителей ВВ и Bb генотипов гена VDR отмечалась более высокая распространенность артериальной гипертонии. Была отмечена связь bb/Aa генотипа с ишемическим инсультом [25].
В арабской популяции El-Barbary AM, et al. обнаружили связь bb генотипа гена VDR и атеросклеротического поражения сосудов [26], в то время как Al-Ghamdi AS, et al. [27] сообщили об отсутствии связи данного генотипа с ИБС в Саудовской Аравии. Небольшое количество обследованных пациентов может частично объяснить противоречивые результаты.
При стратифицированном анализе BsmI полиморфизма гена VDR по этническому признаку было выявлено повышение риска ИБС в европейской популяции у носителей b (A) аллеля на 23% по сравнению с носителями B (G) аллеля. При этом у носителей bb (АА) и Bb (АG) генотипов риск ИБС был на 56% и 20% выше, чем у носителей BB (GG) генотипа [28].
В азиатской популяции не выявлено ассоциации BsmI полиморфизма VDR с риском ИБС. Это может быть связано с тем, что аллели данного полиморфизма расположены между 8-ым интроном и 9-ым экзоном гена VDR, который имеет несбалансированное сцепление в различных этнических группах [28].
ApaI полиморфизм гена VDR влияет на стабильность матриксной РНК [29]. Поскольку VDR играет важную роль в передаче сигналов витамина D, можно предположить, что ApaI полиморфизм гена VDR также может влиять на риск ИБС либо за счет изменения чувствительности рецептора к лигандам, либо за счет ген-генных взаимодействий или влияний окружающей среды на гены.
Носительство Aa (CA) генотипа гена VDR ассоциировалось со снижением риска ИБС у больных СД 2 типа, и, вероятно, данный генотип может быть связан с низким риском ИБС, но для подтверждения этого требуется проведение более крупных исследований [24].
TaqI полиморфизм гена VDR был описан в 1994г [28]. Он обусловлен заменой тимина на цитозин в 352 кодоне. Это приводит к замене кодона AUU (изолейцин) на AUG (метионин) [10]. По результатам метаанализа, tt генотип связан с повышенным риском ИБС в диапазоне от 14% до 27% в различных исследованиях [30].
Результаты исследований связи полиморфизма гена VDR и ИБС противоречивы. Так, например, результаты метаанализа, проведенного Alizadeh S, et al., не показали связи FokI, BsmI, ApaI и TaqI полиморфизмов гена VDR с риском ИБС [30], что может быть связано с различной встречаемостью вышеуказанных генотипов в различных этнических и расовых группах. Например, встречаемость f аллеля (FokI полиморфизм) гена VDR была реже среди африканцев, чем среди азиатов, тогда как В аллель BsmI полиморфизма гена VDR имеет более низкую встречаемость у азиатов, чем у европейцев и африканцев [16]. В литературе практически отсутствуют исследования, посвященные встречаемости FokI, BsmI, ApaI и TaqI полиморфизмов гена VDR и их связи с риском ИБС и особенностями её клинического течения в СевероЗападном регионе Российской Федерации.
Prabhakar P, et al. в своем исследовании не установили связи между носительством различных генотипов ApaI и TaqI полиморфизма гена VDR и развитием ишемического инсульта. У обследованных — носителей ff генотипа гена VDR, риск ишемического инсульта повышался в 2,97 раз, у носителей Ff генотипа — в 1,52 раза. Гомозиготное носительство f аллея гена VDR ассоциировалось с повышением уровня общего холестерина. Носительство bb генотипа гена VDR повышало риск ишемического инсульта в 1,76 раз. Кроме этого, авторами установлено, что лица с дефицитом витамина D (при его уровне в плазме крови <20 мг/мл) также имеют повышенный риск возникновения ишемического инсульта [31].
TaqI, ApaI и BsmI полиморфизмы гена VDR могут влиять на экспрессию мРНК VDR и способствовать снижению уровня витамина D у пациентов с ИБС. Хотя генетические факторы вносят существенный вклад в изменчивость циркулирующих уровней витамина D, при этом наследуемость оценивается в 30-40% [28]. Несбалансированность связей наблюдалась только в данных трех полиморфизмах гена VDR, и они находились в одном гаплоидном домене с 3-регуляторным доменом. Гаплоидный домен включает 4-9 экзонов и 3-регуляторный домен, что связано с регуляцией экспрессии генов, особенно с регуляцией стабильности мРНК.
По-видимому, у носителей прогностически неблагоприятных генотипов гена VDR также отмечается сниженная способность к ингибиции сигнального пути NF-kВ. NF-kВ, в свою очередь, пролонгирует активацию макрофагов и повышает экспрессию адгезивных молекул на эндотелиоцитах, тем самым поддерживая развитие и прогрессирование атеросклеротического поражения сосудов [32].
Эпигенетические механизмы регуляции рецептора витамина D
Результаты исследований последних лет показали, что функционирование 80% генома заключается в эпигенетической регуляции экспрессии генов, кодирующих протеины. МикроРНК представляют собой малые некодирующие РНК, которые являются важнейшими регуляторами экспрессии генов на транскрипционном и посттранскрипционном уровнях. МикроРНК — это одноцепочечные молекулы-регуляторы 30% всех генов. Они имеют специфическую, напоминающую шпильку, структуру, длину от 19 до 24 нуклеотидов, которые образуются из более длинных РНК-предшественников [6]. МикроРНК подавляют экспрессию генов, кодирующих протеины на стадии постранскрипции за счёт неполной гибридизации с 3-регуляторной нетранслируемой областью мРНК, которая имеет комплементарные сайты.
МикроРНК представляют из себя эффективные посттранскрипционные регуляторы генной экспрессии [6]. Они управляют сложными посттранскрипционными регулярными сетями, необходимыми для регуляции экспрессии генов. Таким образом, микроРНК необходимы для тонкой транскрипционной регуляции экспрессии генов, но они также могут играть различные роли в пролиферации, дифференцировке и функционировании определенных типов клеток.
В организме человека в настоящее время выявлено 1 тыс. микроРНК, 50 из которых, вероятно, связаны с риском ССЗ [29].
МикроРНК, как и цитокины, образуют взаимосвязанную регуляторную систему, которая контролирует апоптоз, пролиферацию, дифференцировку тканей. МикроРНК играют важную роль в нормальной физиологии организма, облегчая экспрессию генов в сложных тканевых системах, но и имеют значение в развитии патологических состояний, включая дисфункцию эндотелия, формирование с последующим разрывом атеросклеротической бляшки [33, 34]. Некоторые микроРНК рассматривают как возможные диагностические маркеры ИБС [6][34].
Экспрессионная активность гена VDR может модулироваться как самим витамином D, так и другими факторами, играющими важную роль в эпигенетических модификациях, такими как микроРНК [35].
Регуляторные сети микроРНК особенно важны для сигнальных молекул, к которым относится витамин D, оказывающих плейотропное действие на различные органы и ткани организма. В недавних исследованиях показана роль 1,25(OH)D в эпигенетической регуляции генов, особенно в качестве модулятора функции микроРНК [36].
Показано, что экспрессия VDR коррелирует с экспрессией таких провоспалительных цитокинов, как интерлейкин-1, 6 и 8, и может регулироваться микроРНК [36]. Авторы объясняют данную корреляцию тем, что микроРНК могут играть решающую роль в связанных с ожирением изменениях экспрессии VDR посредством местного влияния витамина D в развитии локального воспаления в жировой ткани, однако для подтверждения этого наблюдения необходимы дальнейшие исследования [37].
Как уже ранее упоминалось, витамин D подавляет экспрессию проангиогенной микроРНК-155, которая представляет из себя отрицательный регулятор экспрессии супрессора передачи сигналов цитокинов, путем ингибирования патогенетического пути NF-?B [35].
Потенциальный сайт связывания микроРНК-125b был идентифицирован в 3-нетранслируемой области гена VDR человека [38], где локализованы BsmI, ApaI и TaqI полиморфизмы гена VDR, которые потенциально могут оказывать влияние на взаимодействие микроРНК-125b и гена VDR. В исследовании на культуре клеток MCF-7 (эпителиоподобная клеточная линия, полученная из инвазивной аденокарциномы протоков молочной железы человека) установлено, что повышенная экспрессия микроРНК-125b сопровождалась снижением уровня эндогенного белка VDR на 40% [38].
Экспрессия VDR в висцеральной жировой ткани у пациентов с ожирением отрицательно коррелировала с уровнями микроРНК-125a-5p, микроРНК125b-5p и микроРНК-214-3p [36].
Сайт связывания микроРНК-21 также расположен в 3-регуляторном домене гена VDR [38] и, как и в случае с микроРНК-125b, потенциально может подвергаться влиянию одиночных нуклеотидных полиморфизмов гена VDR, локализованных в данной области. МикроРНК-21 экспрессируется в эндотелиоцитах и регулирует их функции. Результаты исследований, посвященных поиску роли данной микроРНК в атерогенезе, противоречивы. Lisse TS, et al. в 2013г установили, что при увеличении экспрессии микроРНК-21 в эндотелиоцитах снижается их миграция и пролиферация, что может свидетельствовать об антиангиогенной функции данной микроРНК. Экспрессия микроРНК-21 существенно повышена в атеросклеротических бляшках и макрофагах [30]. Следовательно, повышенная экспрессия микроРНК-21 может указывать на прогрессирующий рост атеросклеротических бляшек.
МикроРНК взаимодействуют с 3-нетранслируемым регионом таргетной мРНК посредством комплиментарности с её последовательностями, и в первую очередь подавляют экспрессию генов, вызывая деградацию или ингибирование трансляции транскриптов посредством частичного комплементарного связывания пар оснований с тремя первичными нетранслируемыми или экзонными областями. На функционирование микроРНК также могут повлиять и другие факторы, прежде всего изменения структуры генов — генетические полиморфизмы, амплификация, делеция генов, нарушение процессинга и метилирование [37].
VDR может либо подавлять, либо индуцировать микроРНК посредством двух механизмов:
1) прямой регуляции транскрипции через промотерные последовательности гена VDR или гена микроРНК;
2) косвенной регуляции с вовлечением других транскрипционных факторов.
МикроРНК, в свою очередь, могут регулировать действие, синтез и метаболизм витамина D или сами подвергаться влиянию сигналов VDR благодаря механизмам динамической обратной связи, что может приводить к дестабилизации мРНК и/или к ингибиции трансляции [30].
Заключение
Таким образом, в последние годы активно исследуется роль генетического полиморфизма гена VDR и микроРНК в развитии ИБС и её осложнений. Результаты исследований связи полиморфизма гена VDR и ИБС противоречивы, что может быть связано с различной встречаемостью вышеуказанных генотипов в различных этнических и расовых группах. В литературе практически отсутствуют исследования, посвященные встречаемости FokI, BsmI, ApaI и TaqI полиморфизмов гена VDR и их связи с риском ИБС и особенностями её клинического течения в СевероЗападном регионе Российской Федерации.
Экспрессия VDR может регулироваться микроРНК125а-5р, микроРНК-125b-5р, микроРНК-214-3р и микроРНК-21 и иметь отрицательную корреляционную связь с уровнем провоспалительных цитокинов, однако для подтверждения этих данных требуется проведение более крупных исследований. В литературе имеются единичные работы, показавшие, что вышеуказанные микроРНК сами могут подвергаться влиянию полиморфных вариантов гена VDR благодаря механизмам динамической обратной связи, что может приводить к дестабилизации мРНК и к ингибиции трансляции. Поэтому исследование генетических и эпигенетических факторов, регулирующих экспрессию и функционирование VDR, крайне актуально.